Введение в ксенобиологию, часть вторая
Начало
Окончание
2. Химия биологии
Если мы пока решим ограничиться только областью низких энергий, где существуют химические соединения, то прежде всего нас будет интересовать аналог самокопирования в химии - автокаталитические реакции. Реакции эти довольно распространены и аналоги их можно подобрать и в ядерной физике, но к жизни они не имеют отношения до тех пор, пока при автокатализе не происходит образование структур такой сложности, одномоментное самовозникновение которых в данной среде исчезающе маловероятно. Но следует заметить, что возникновение жизни вообще все таки может быть не столь уж пессимистично-маловероятным (как писал Е.Кунин в «Логике Случая»), особенно в свете того, что эволюционный потенциал переходного - предбиологического - этапа нам очень мало понятен. Тут вполне могут быть полезными, например, давно известные свойства палиндромных последовательностей.
Что же касается альтернативной биохимии, то тут, несмотря на очень веские доводы в пользу водно-углеродного шовинизма, тщательнейше собранные и изложенные Михаилом Никитиным, некоторые другие «экзотические» варианты, среди которых наиболее часто можно встретить описание организмов со значительным участием кремния в скелете главных биомолекул или же аммиака в качестве биорастворителя вместо воды - всё ещё не могут быть полностью исключены, но скорее всего их эволюционный потенциал и вероятность возникновения заметно меньше, чем у нашей. А накладываемые химией ограничения и минимальные прямые аналогии позволяют нам представить хотя бы общие очертания этих возможностей.
Почти все теоретики согласны с тем, что главных химических «ингридиентов» жизни должно быть как минимум два сорта: те, из которых строятся сами биомолекулы и те, которые образуют среду для этого строительства. Первые должны быть способны образовывать большие, сложные и в то же время достаточно стойкие молекулы, а вторые - обеспечивать их хорошее перемешивание (диффузию) в равномерных условиях, что возможно только в жидкостях.
В качестве структурообразующих элементов этим условиям удовлетворяют разве что бор, углерод, кремний и фосфор. Но бора при термоядерном синтезе в естественных условиях образуется очень мало, фосфора - побольше, но цепи из него малостабильны (причем это свойство тоже запользовано нашей жизнью как оперативное хранилище химической энергии в виде АТФ), а кремний не обладает способностью образовывать сложные пи-связи как углерод, поэтому «чисто-кремниевая» (полисилановая) жизнь не возможна - совсем без углерода обойтись не удастся и максимум возможного здесь - базис из кремний-органических молекул: органосилоксанов и/или органосилатранов. Тем более если мы обратимся к составляющим нас якобы «углеродным» главным деталям - азотистым осонованиям нуклеиновых кислот и пептидам - то там мы увидим похожую картину - в их скелете (центральной цепи атомов) практически на каждые два атома углерода приходится один атом азота:
Рис.1. Природные азотистые основания ДНК (A) и фрагмент пептидной цепи (B).
Зеленый цвет - атомы углерода, синий - азота, красный - кислорода
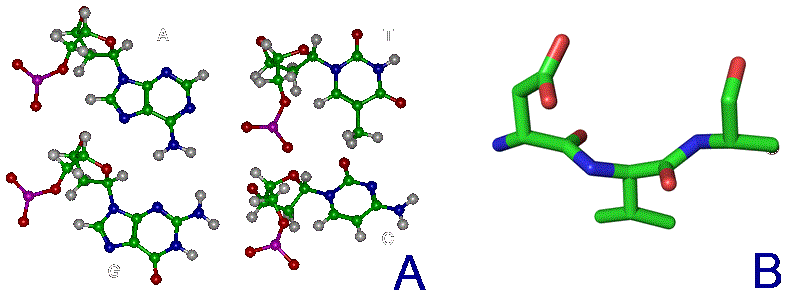
Так что с этой точки зрения куда правильнее было бы называть нашу жизнь «углерод-азотной». И ещё не следует забывать про большое количество кислорода в углеводах и прочие элементы - от фосфора (по одному атому на нуклеотид) и серы (хотя бы пара атомов есть практически в любой более-менее сложной белковой молекуле) и вплоть до редкого молибдена, без которого эффективный механизм фиксации вышеупомянутого азота из атмосферы для постройки своего тела эволюция так и не смогла создать. Еще одно ограничение, накладываемое на кремниевую жизнь состоит в том, что синтезу кремний-органики довольно сильно препятствует наличие воды, а это в целом гораздо более распространенное соединение во Вселенной, чем оксид кремния.
Все это находится в полном соответствии с приведенным выше «математическим» определением жизни - согласно ему в основе жизни должны лежать довольно стабильные и достаточно точно копирующиеся нерегулярные последовательности конечного числа блоков, а вот скелет цепи этих блоков должны составлять уже регулярные чередования разных атомов: хотя достаточно длинные однородные, «гомоатомные» цепи и могут существовать (пример - нонаконтатриктан), операции с ними (синтез, обратимое разделение и сшивка) «подручными средствами» (без широкого изменения физических параметров среды) крайне затруднены.
Для упрощения понимания можно провести такую аналогию: наши методы химического синтеза работают подобно заводским пресс-формам или как минимум 3D-принтеру - довольно большому и сложному механизму, производящему монолитные вещи, тогда как биополимеры гораздо более напоминают LEGO-конструктор: если у вас есть только один тип блоков - вы многого сделать не сможете, а чтобы блоки можно было легко перекомбинировать, сами они должны иметь неоднородность, но повторяющуюся (паз-выступ). Регулярные цепи из разных элементов - это и есть полимеры.
Аналогию с LEGO можно даже углубить: всякий, кто пытался собрать что-то действительно сложное из такого конструктора, знает как ценны в наборе детали, имеющие боковые выступы (так называемые SNOT-элементы).
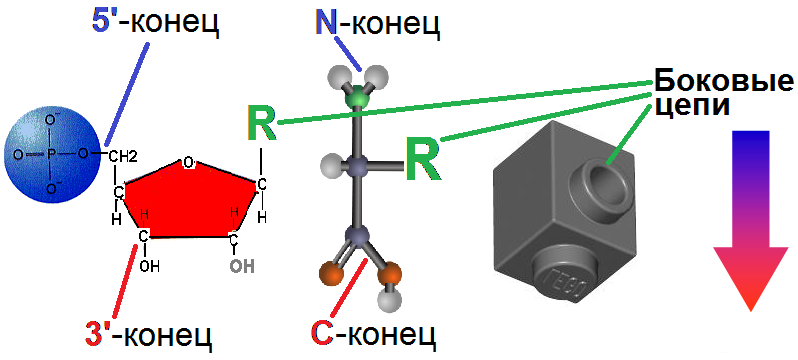
Совершенно очевидно каждому: как повышает разнообразие конструкций «свобода маневра» в трех измерениях. Элементарные детали обоих наших биополимеров - нуклеиновых кислот и белков - нуклеотиды и аминокислоты - устроены именно как SNOT-элементы LEGO, хотя их боковые цепи не позволяют ветвить скелет (впрочем, и в реальных конструкторах эти связи прочностью обычно не отличаются):
Рис.2. Элементарные звенья известных биополимеров.
Слева направо: нуклеотид, аминокислота, SNOT-элемент. Стрелкой показано направление роста цепей.
Отсюда же и произрастает явление зеркальной несимметричности (хиральности) биомолекул: боковые ветви симметричной цепи дают разнообразие только в двух измерениях. При этом наши биополимеры обладают тут интересной особенностью: нуклеиновые кислоты и белки живых организмов состоят только из мономеров одной из зеркально-симметричных форм, тогда как при обычном химическом синтезе обе формы образуется практически в равных соотношениях - это так называемая проблема хиральной чистоты биополимеров(homochirality). Биологическое значение этой избирательности очевидно: такая унификация сильно упрощает обеспечение пространственной (третичной) структуры этих макромолекул, от чего прямо зависит их функция.
Кстати: так легко объясняется загадочная на первый взгляд спиральная упорядоченность многих биологических объектов начиная с ДНК: это просто многократно повторенная в одну и ту же сторону асимметричность составляющих её элементов, причём усложняющая ей жизнь (расплетение этой спирали при копировании (репликации) требует специальных ферментов - топоизомераз), поскольку никакой более сложной пространственной (третичной) структуры ДНК не образует, что ей, как пассивному хранилищу информации, и не требуется. Но требовалось её прямому предшественнику - РНК в эпоху «РНК-мира», когда один тип молекул нес на себе функции и хранения и реализации наследственности . РНК же третичную структуру очень даже имеет, активно формируя связи не только между двумя отдельными, полностью комплементарными цепями, но и в пределах одной цепи, также как и белки образуя сложные трехмерные «загогулины», что было бы невозможно, если бы составляющие её звенья-нуклеотиды были симметричными.
Это наглядно видно даже в структуре самых маленьких цепочек - транспортных РНК наших клеток:
Рис.3. Свернутая в трехмерном пространстве(третичная) и «расплющенная» на плоскости(вторичная) структуры молекулы транспортной РНК.
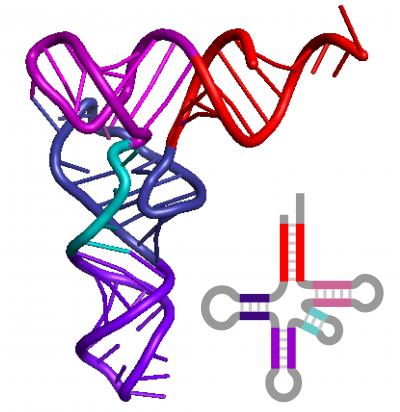
Таким образом «завораживающую спиральность ДНК» можно рассматривать просто как наследие (рудимент) РНК-мира.
Механизмы «очистки» исходной смеси изомеров теперь понемногу проясняются, но первопричина и вероятность выбора жизнью той или иной формы - по сей день остаются не до конца ясными. Можно сказать, что тут существует два подхода:
1) «Нулевая гипотеза» - первая молекула РНК-репликатор должна иметь либо правую, либо левую хиральность, но произошло это чисто случайно (как подброшенная монета не упадет на ребро) - а дальше заработала биологическая эволюция по принципу «кто первый встал - того и тапки».
2) Гипотеза Вестера-Ульбрихта, согласно которой гомохиральность жизни является следствием фундаментальной пространственной несимметричности слабого взаимодействия элементарных частиц.
Буквально во время написания данного текста была опубликована работа, которая хотя и не снимает, но очень изящно ставит эту «проблему» не в начало абиогенеза, а в его середину и на службу куда более важной задаче - созданию первой самореплицирующейся молекулы РНК:рибозимы-полимеразы гораздо эффективнее копируют зеркальные варианты друг друга, чем свои собственные! Причём длина цепи нужной для этого молекулы РНК оказывается более чем в два раза короче, чем в случае всех предыдущих подходов - около 83 нуклеотидов (как у вышепоказанной транспортной РНК), тогда как в «Логике Случая» в качестве ориентировочного максимального предела «неуменьшаемой (нередуцируемой) сложности», который может быть собран в нашей Вселенной просто случайным перебором нуклеотидов с вероятностью, не превышающей количество имеющихся ресурсов и времени Е.Куниным была установлена уникальная последовательность длиной 102 нуклеотида, причем в случае рассматриваемой работы очевидно, что это - не единственный и даже не самый эффективный вариант работающей последовательности!
Более того: околотопологические подходы (по сути - «передний край» современной математики) к проблемам хиральности, узлов, фракталов и пр. в применении к задаче построения самокопирующихся молекулярных машин вообще оказываются неожиданно продуктивными.
Таким образом сложная химическая жизнь возможна только на базе нерегулярных хиральных гетерополимеров.
Сложные машины из конструктора с правильным набором деталей подручными средствами или даже путем случайных перемешиваний сделать гораздо проще, чем из большого количества простых, даже высококачественных, но никак не обработанных материалов. Известная метафора креационистов о невозможности урагана на свалке собрать из мусора Боинг-747 неадекватна еще и по этой причине. Чтобы приблизить её к реальности лучше говорить о торнадо, переворачивающем склад SNOT-элементов фабрики LEGO, причем на регулярной основе - а в этом случае результат уже не так однозначен. Особенно если эти SNOT-блоки снабдить сильными магнитами.
Другой часто описываемый вариант - замена биорастворителя - воды на аммиак - ограничена не только заметно более узкой «зоной жизни» для гипотетических планет с аммиачными океанами вследствие того, что жидкий аммиак существует при меньшем диапазоне температур (при стандартном давлении - вдвое по сравнению с водой), но и таким фактором, как жесткий ультрафиолет: УФ-расщепление аммиака (фотолиз) может быть обратимым только на достаточно крупных планетах, способных удерживать в атмосфере водород.
Кроме того некоторые исследователи возлагают определенные надежды на холодные планеты с углеводородными океанами, как на Титане, что встречает препятствие иного рода: в сравнении с аммиаком или водой углеводороды - практически неполярные молекулы - то есть электронная плотность в них распределена довольно равномерно: «более отрицательно» или «более положительно» заряженных мест в них не выделяется.
Неполярные же молекулы хорошо растворяются только в неполярных растворителях, но практически нерастворимы в полярных (и наоборот). И на этих взаимодействиях и с учетом их построены все структуры в клетках - на этом сочетании базируется почти вся молекулярная самоорганизация и прежде всего - существование клеточной мембраны.
В нашем случае мембрана обычно построена из фосфолипидов, молекулы которых состоят из больших неполярных углеводородных «хвостов» и полярной «головки»: в полярном растворителе двойной слой молекул ориентирован «хвостами» друг к другу, а «головами» - к растворителю (рис. 4A).
Можно, например, попытаться представить и «клетку наизнанку» - с неполярным растворителем в цитоплазме и окружающей среде, разделяемым в основном полярной мембраной (рис. 4B), но нужно понимать, что полярные молекулы притягиваются между собой заметно сильнее, чем неполярные за счет водородных связей - именно поэтому у воды, аммиака и полярной органики (многоатомных спиртов, карбоновых кислот, углеводов и др.) выше температуры плавления и кипения и шире диапазон жидкого состояния.
Рис.4. Обычная фосфолипидная (A) и гипотетическая «инвертированная» (B) мембраны.
Желтый цвет - неполярные("жирные") слои, голубой - полярные("водные") слои.

Грубо говоря: свойства таких мембран будут отличаться как свойства капли масла в воде и капли воды в масле, причем многократно усиленные на микромасштабах.
Само же наличие гибкой полупроницаемой оболочки, по всей видимости, необходимо для появления любых хоть сколь-нибудь сложных организмов (сложнее вирусов), иначе мы скорее всего снова встанем перед проблемой нередуцируемой сложности, когда жесткая оболочка потребует сразу слишком большое число транспортных систем для обмена всеми нужными разновидностями веществ со внешней средой.
Причем те автономные организмы, которые некогда пошли по пути образования малопроницаемых оболочек, хотя и существенно повышают устойчивость во внешней среде, в то же время на несколько порядков теряют в скорости роста и размножения и в результате имеют довольно примитивное строение. Наглядный пример: наши микобактерии (в их числе - возбудители туберкулеза и лепры).